
jueves, 12 de abril de 2018
lunes, 9 de abril de 2018
viernes, 16 de marzo de 2018
Sistema Endócrino
El sistema endocrino está formado por neuronas y glándulas que producen y secretan hormonas, que son sustancias químicas producidas en el cuerpo que regulan la actividad de las células y los órganos. Estas hormonas regulan el crecimiento del cuerpo, el metabolismo (los procesos físicos y químicos del cuerpo) y el desarrollo y la función sexual. Las hormonas se liberan en el torrente sanguíneo y pueden afectar uno o varios órganos en todo el cuerpo.
Las hormonas son mensajeros químicos creados por el cuerpo. Transfieren información de un conjunto de células a otro para coordinar las funciones de las diferentes partes del cuerpo.
as glándulas pueden ser los dos tipos siguientes:
Las hormonas son mensajeros químicos creados por el cuerpo. Transfieren información de un conjunto de células a otro para coordinar las funciones de las diferentes partes del cuerpo.
as glándulas pueden ser los dos tipos siguientes:
- Glándulas exocrinas, que segregan sus productos mediante conductos (glándulas digestivas, sudoríparas, mamarias).
- Glándulas endocrinas, que segregan las hormonas en la sangre y llegan a actuar sobre órganos o tejidos diana.
Objetivos del blog
Este blog forma parte de una práctica escolar de la materia de Nuevas Tecnologías de la Información y la Comunicación de la Universidad de Sonora
 Actualmente curso la Licenciatura en Enfermería, y he tomado como objetivo propio el usar este blog como apoyo para mis estudios en la anatomía y fisiología del cuerpo humano, plasmando y estructurando en él toda información que considero importante.
Actualmente curso la Licenciatura en Enfermería, y he tomado como objetivo propio el usar este blog como apoyo para mis estudios en la anatomía y fisiología del cuerpo humano, plasmando y estructurando en él toda información que considero importante.

Hormonas adenohipofisiarias. Formación y sus efectos
Hormona tiroestimulante o tirotrofina (TSH).
A nivel hipotalámico se sintetiza TRH (u hormona liberadora de tirotropinag); esta hormona estimula la síntesis y liberación de TSH por la adenohipófisis. La concentración de TSH estimula la actividad de la bomba de yoduro en las células tiroideas, regulando así la cantidad de T3 y T4 que se genera. Para la formación de las hormonas tiroideas se debe oxidar el yodo. Esto depende de la enzima perioxidasa y el peróxido de hidrógeno que lo acompaña. Éstos se encuentran justo donde la molécula de tiroglobulina será secretada, para que el yodo oxidado, se pueda ya unir directamente a la tirosina y atravesar la membrana hasta donde el coloide será almacenado. La tiroxina, que es el principal producto hormonal, se forma cuando una tirosina se yoda primero a Monoyodotirosina, después a Diyodotirosina y al final 2 moléculas de Diyodotirosina se unen haciendo T4. La triyodotirosina T3 se forma cuando una monoyodotirosina se une con una diyodotirosina o cuando una tiroxina pierde un yoduro.La hormona tiroidea se segrega gracias al yodo que se ingiere con los alimentos (y de la sal*). este yodo llega a la sangre (yodemia) y parte de él se pierde por la orina. El resto, cuando llega a la glándula tiroides pasa a ser yodo orgánico gracias a la acción de la peroxidasa. Este yodo orgánico + tirosina (aminoácido) forman monoyodotirosina. Y consecuentemente;– Monoyodotirosina + Monoyodotirosina = Diyodotirosina
– Diyodotirosina + Monoyodotirosina = Triyodotirosina (T3)
– Triyodotirosina + monoyodotirosina = Tiroxina (T4)
T3 y T4 circulan unidas a proteínas y se destruyen en los músculos y en el hígado.
Las hormonas tiroideas son liposolubles y pueden atravesar la membrana plasmática de sus células diana por difusión o por un proceso mediado por transportador. Una vez en el citoplasma T4 se transforma en T3, de modo que los niveles citoplasmáticos de T3 y T4 son similares, y se unen a un gran número de lugares. Tanto T3 como T4 entran en el núcleo donde están sus receptores que, por tanto, son receptores nucleares (ver el apartado de endocrinologia, generalidades). Los receptores de hormonas tiroideas se unen al ADN en la región promotora de genes regulados por dichas hormonas, de modo que la unión de T3 y T4 a sus receptores promueve la transcripción de un gran número de genes codificadores de un amplio rango de proteínas. Debido a ésto, los efectos de las hormonas tiroideas suelen tardar en aparecer varios días después de la estimulación de la secreción de la glándula tiroides. Hay un aumento del ARN mensajero seguido por un aumento de la síntesis de proteínas que genera un incremento en los niveles intracelulares de enzimas específicos. Por tanto en la mayor parte de los tejidos aumenta el número de enzimas, proteínas estructurales, proteínas de transporte y otras sustancias. El resultado es un incremento generalizado de la actividad funcional de todo el organismo. Efectos sobre las proteínas: a concentraciones normales, la T3 y T4 estimulan la captación de aminoácidos en las células y la síntesis de proteínas estructurales y funcionales específicas. Efectos sobre los lípidos: la T3 y T4 tienen un efecto lipolítico sobre los depósitos de grasa del organismo con lo que aumentan los niveles de ácidos grasos libres en el plasma. También producen un aumento de la oxidación de los ácidos grasos libres lo que contribuye al efecto productor de calor que tienen estas hormonas. Efectos sobre los hidratos de carbono: la T3 y T4 aumentan la absorción intestinal de glucosa y la captación de la misma por las células del organismo, sobre todo las musculares y adiposas. Facilitan la gluconeogénesis porque aumentan la disponibilidad de los materiales necesarios (aminoácidos y glicerol), actúan directamente sobre los enzimas implicados en la glicolisis, activándolos, y potencian de un modo indirecto la acción sobre los hidratos de carbono de otras hormonas como la insulina y las catecolaminas.
HORMONAS TIROIDEAS T3 Y T4. EFECTOS EN LA PRODUCCIÓN DE CALOR Y
EL CONSUMO DE OXÍGENO
En la mayor parte de los tejidos, la T3 y T4 aumentan la producción de calor y el
consumo de oxígeno.
La hormona adrenocorticotropa, corticotropina o corticotropina (ACTH).
La liberación de ACTH (corticotropina) por la hipófisis depende de la hormona hipofisiotrópica CRH. Una vez que se secreta ACTH a la sangre, esta tiene un rápido efecto sobre las dos zonas profundas de la corteza suprarrenal especialmente la zona fascicular, para aumentar la producción de cortisol. Este efecto de la ACTH se consigue mediante el aumento de la conversión de colesterol en pregnenolona, y está mediado por el segundo mensajero AMPcíclico. Las concentraciones sanguíneas de cortisol libre (no unido) se controlan por un mecanismo de retroacción. Unas concentraciones plasmáticas de cortisol mayores reducen la secreción de ACTH mediante un efecto directo sobre la hipófisis, así como una inhibición indirecta de la liberación de CRH por el hipotálamo. La secreción de cortisol es máxima a primeras horas de la mañana y alcanza su mínimo a últimas horas de la tarde. La esteroidogénesis suprarrenal es la entrada de colesterol a través de las membranas mitocondriales externa e interna. En la membrana mitocondrial interna, se escinde la cadena lateral de colesterol para producir pregnenolona. Esta reacción es catalizada por la enzima divisora de la cadena lateral del colesterol (colesterol desmolasa, P450scc, CYP11A1), una enzima del citocromo P450 (CYP). Los mecanismos por los que se regulan los andrógenos suprarrenales, DHEA y androsteidona, no están completamente aclarados. Mientras que la ACTH estimula la producción de
andrógenos suprarrenales de forma aguda y es el principal estímulo de la liberación de cortisol, se han identificado factores adicionales
La Glucocorticoides sobre el metabolismo de los hidratos de carbono es el aumento de la producción de glucosa mediante el aumento de la gluconeogénesis hepática. También aumentan la resistencia celular a la insulina, por lo que disminuyen la entrada de glucosa a la célula.
El MC más importante es la aldosterona y en menor grado la 11-desoxicorticosterona. Los MC tienen acciones más limitadas que los GCC. Su función fundamental es mantener el volumen intravascular conservando sodio y eliminando potasio e hidrogeniones. Ejercen estas acciones en el riñón, el intestino y las glándulas salivales y sudoríparas.
Muchas acciones de los andrógenos suprarrenales se ejercen mediante su conversión a andrógenos o estrógenos activos, como la testosterona, la dihidrotestosterona, la estrona y el estradiol.
Todas ellas son sintetizadas a partir del esteroide colesterol y tienen fórmulas químicas similares. El colesterol es captado por las células glandulares de un modo directo desde la sangre porque la membrana de estas células tiene receptores en donde se fijan las lipoproteínas de baja densidad (LDL) que transportan concentraciones elevadas de colesterol. Al fijarse estas lipoproteínas a los receptores de la membrana celular se fomenta la entrada de colesterol en la célula por un mecanismo de endocitosis mediada por receptor. Las células glandulares también pueden sintetizar colesterol a partir del Acetil-CoA, pero este proceso se da en menor proporción. El colesterol es luego almacenado dentro de gotas de lípidos
en el citoplasma de las células de la corteza suprarrenal.
El cortisol se une a un receptor citoplasmático y el complejo hormona-receptor entra en el núcleo celular y modula la trascripción de genes en muchos tejidos. El cortisol es esencial para la vida. El 95% de la actividad glucocorticoide suprarrenal se debe al cortisol o hidrocortisona. En menor proporción intervienen otros glucocorticoides. Junto con otros glucocorticoides realiza numerosas acciones en todo el organismo
en donde desempeña un papel fundamental en la respuesta del organismo al estrés, tanto físico como emocional. Es antiinflamatorio, ya que evita los efectos dañinos de la reacción inflamatoria al
disminuir la actividad de las células que participan en las reacciones inflamatorias. Reduce el número de mastocitos y, por tanto, la liberación de histamina y otras sustancias vasoactivas con lo que la permeabilidad capilar es menor. Por lo mismo, en caso de alergia, modifica la respuesta inflamatoria provocada por la reacción antígeno-anticuerpo, que se hace menos intensa. Es inmunosupresor ya que disminuye el número de linfocitos T y B circulantes, con una menor producción de anticuerpos. Actúa sobre el sistema nervioso central produciendo euforia y otros cambios de humor. Aumenta el tono vascular, posiblemente al potenciar los efectos de las catecolaminas y hace a los vasos sanguíneos más sensibles a los vasoconstrictores
con lo que contribuye a elevar la presión arterial.
CORTISOL. EFECTOS METABÓLICOS
Las principales acciones de esta hormona consisten en regular el metabolismo de la glucosa, disminuyendo su utilización en los tejidos periféricos y aumentando la gluconeogénesis y los niveles de glicemia. Es una hormona fundamental en la resistencia del organismo al estrés. Efectos sobre las proteínas. Disminuye la síntesis de proteínas en el organismo, con excepción del hígado. Aumenta el catabolismo de las proteínas y el traslado de los aminoácidos desde las células, sobre todo las fibras musculares, hasta el hígado, en donde los aminoácidos pueden ser convertidos en nuevas proteínas como los enzimas que son necesarios para las reacciones metabólicas o las proteínas de la coagulación. Efectos sobre los lípidos. Estimula la lipólisis, es decir, la ruptura de los triglicéridos en ácidos grasos y glicerol, y la liberación de los ácidos grasos del
tejido adiposo al plasma. Efectos sobre los hidratos de carbono. Disminuye la captación y utilización de
glucosa por las células con lo que aumentan los niveles de glucosa en plasma (glicemia). Causa una rápida movilización de los aminoácidos y de las grasas de sus lugares de depósito (músculo y tejido adiposo, respectivamente) dejándolos disponibles para obtener energía de ellos en vez de la glucosa
Los efectos de los mineralcorticoides son esenciales para la vida, de modo que si no hay, se produce la muerte al cabo de pocos días. La función más importante de la aldosterona es la reabsorción de sodio a nivel de la parte distal de las nefronas, en los riñones, con lo que el sodio se recupera a la sangre y no se pierde en la orina. Este efecto se realiza mediante un intercambio con potasio que se elimina por la orina y no se acumula en el organismo. La reabsorción de sodio conduce a la reabsorción de los iones cloro y bicarbonato y al mantenimiento de agua en el organismo. Por tanto, la aldosterona evita la depleción de sodio del organismo, ya que conserva sodio en el líquido extracelular, y controla los niveles extracelulares
de potasio (fundamentales para el buen funcionamiento celular) evitando una acumulación de este ión. Si hay una disminución en la secreción de aldosterona se produce un aumento en los niveles de potasio en plasma (muy peligroso para la vida) y una disminución de los de sodio y cloro. Como consecuencia, el volumen de
líquido extracelular y el volumen sanguíneo disminuyen y se produce una disminución del gasto cardíaco que puede llevar a la muerte. Debido a su solubilidad en los lípidos de la membrana, la aldosterona difunde con
facilidad hacia el interior de las células epiteliales de los túbulos de las nefronas (distales y colectores) en los riñones. En el citoplasma de estas células se combina con un receptor citoplasmático. El complejo receptor-hormona difunde al núcleo donde da lugar a la transcripción de genes y formación de RNA mensajero. Este
RNA mensajero difunde al citoplasma y origina la formación de una o más proteínas transportadoras de sodio y de potasio.
La cantidad de hormonas sexuales secretadas por la corteza suprarrenal de un adulto normal es tan baja que sus efectos son insignificantes. En la mujer, los andrógenos suprarrenales contribuyen a la libido. Asimismo, los andrógenos suprarrenales contribuyen al crecimiento prepuberal y al desarrollo del pelo axilar y púbico, tanto en chicas como en chicos.
El exceso de cortisol puede producir algunos de los signos distintivos del síndrome de Cushing: una joroba de grasa entre los hombros, la cara redondeada y estrías gravídicas de color rosa o púrpura en la piel. El síndrome de Cushing también puede derivar en presión arterial alta, pérdida ósea y, a veces, diabetes tipo 2.
Sus efectos negativos se perciben con el esceso de cortisol y son las elevación de exagerada de azúcar en sangre, el aumento de la presión arterial, osteoporosis, la producción excesiva de vello, la generación de estrías en la piel del abdomen, retención de agua en los párpados, la cara de luna llena, la debilidad muscular, obesidad del tronco y el agotamiento. Además de enfermedades tales como:
1) Síndrome de Addison el que se genera con la falta de cortisol.Los síntomas de la enfermedad de Addison se suelen desarrollar lentamente, a menudo, durante varios meses, y pueden incluir lo siguiente:
Fatiga extrema
Pérdida de peso y disminución del apetito
Oscurecimiento de la piel (hiperpigmentación)
Presión arterial baja, incluso desmayos
Ansias de consumir sal
Bajo nivel de azúcar en sangre (hipoglucemia)
Náuseas, diarrea o vómitos
Dolor abdominal
Dolores musculares o de las articulaciones
Irritabilidad
Depresión
Caída del vello corporal o disfunción sexual en las mujeres
2) Síndrome de Cushing el que se ocasiona con el aumento de cortisol.Los síntomas de la enfermedad de Cushing incluyen:
Obesidad de la parte superior del cuerpo (por encima de la cintura), y brazos y piernas delgados
Cara redonda, roja y llena (cara de luna llena)
Tasa de crecimiento lenta en los niños
Los cambios en la piel que se ven con frecuencia incluyen:
Acné o infecciones de la piel
Marcas de estiramiento purpúreas (1/2 pulgada o 1 centímetro o más de ancho), llamadas estrías en la piel del abdomen, los muslos, la parte superior de los brazos y las mamas
Piel delgada con propensión a la formación de hematomas, sobre todo en los brazos y las manos
Los cambios en los músculos y los huesos incluyen:
Dolor de espalda, que ocurre con las actividades rutinarias
Dolor o sensibilidad en los huesos
Acumulación de grasa entre los hombros (joroba de búfalo)
Debilitamiento de los huesos, lo cual lleva a fracturas en las costillas y la columna vertebral
Músculos débiles
Las mujeres pueden tener:
Crecimiento excesivo de vello en la cara, el cuello, el pecho, el abdomen y los muslos
Ciclo menstrual que se vuelve irregular o cesa
Los hombres pueden tener:
Disminución o ausencia del deseo sexual (baja libido)
Problemas de erección
Hormona foliculoestimulante (FSH).
La síntesis y la liberación de las hormonas gonadotrópicas
hipofisarias, son reguladas por la hormona hipotalámica liberadora de gonadotrofinas (GnRH). La hormona GnRH, se libera en forma de pulsos, los cuales pueden ser regulados por señales externas al hipotálamo, tales como las hormonas esteroideas.
La baja frecuencia de pulsos de GnRH conduce a la liberación de FSH, mientras que la alta frecuencia de pulsos de GnRH estimula la liberación de LH. La GnRH es excretada en las venas portales de la pituitaria para su transporte a las células gonadotróficas de la pituitaria anterior donde se une a los receptores de la membrana. Una vez fijada a sus receptores la GnRH provoca una entrada de calcio que se actúa como segundo mensajero uniéndose a la calmodulina y haciendo que las células liberen simultáneamente hormona luteínica (LH) y hormona estimulante del folículo (FSH) de sus gránulos secretores. La GnRH también actúa estimulante la transcripción de los genes que dirigen la síntesis de las dos gonadotropinas.
El cuerpo lúteo también conocido como cuerpo amarillo es una masa amarilla formada a partir del folículo (se forma cuando el óvulo sale del folículo y este folículo se transforma en cuerpo amarillo) aparece en el ovario después de la ovulación, donde la cual se regenera una serie de hormonas , en concreto estrógeno y progesterona.El cuerpo lúteo es una glándula endocrina temporal originada por las células foliculares y de la teca interna remanentes de un folículo post ovulación. Esta glándula secreta principalmente progesterona. Luego, la progesterona actuará sobre la mucosa uterina, transformándola en un tejido secretor que luego albergará el huevo fecundado o embrión. El folículo es una estructura anatómico-funcional que forma parte del ovario y el ovocito es la célula que irá madurando en una parte microscópica de pared interna de un folículo a lo largo del ciclo ovárico espontáneo o estimulado en condiciones normales. Además, el folículo contiene otro tipo de células que producen estrógenos, necesarios para el normal desarrollo de la maduración de los óvulos. El foliculo ovarico es el que produce los ovulos, y el cuerpo luteo se forma despues que el ovulo sale del foliculo, la diferencia seria que el foliculo se trasnforma en cuerpo luteo
En el hombre, una insuficiencia testicular denominada hipogonadismo.
En la mujer, la medición de los valores de esta hormona permite diagnosticar una amenorrea, esterilidad o hipogonadismo.-Infertilidad.-Quistes ováricos.
Hormona luteinizante
(LH).
En las mujeres, un aumento de la LH desencadena la ovulación. Influye en el incremento de la actividad aromatasa en las células de la granulosa y en la producción de andrógenos en las células de la teca durante la maduración folicular preovulatoria. En el momento de la maduración del folículo, el aumento de estrógenos conduce a una liberación de LH que desencadena la ovulación, no sólo liberando el óvulo sino también iniciando la conversión de los folículos residuales en un cuerpo lúteo que, a su vez, produce progesterona con el fin de preparar al endometrio para una posible implantación. En el hombre, la LH actúa sobre las células de Leydig del testículo y se encarga de la producción de testosterona, un andrógeno que ejerce actividad endocrina y actividad intratesticular (como la espermatogénesis).
Hiposecreción: En hombres, se observa una producción insuficiente de espermatozoides. En mujeres, se manifiesta como amenorrea. Hipersecreción: Menopausia prematura, castración, insuficiencia testicular, síndrome de ovario poliquístico, síndrome de Klinefelter, síndrome de Swyer, síndrome de Turner.
Prolactina (PRL).
En contraste con las demás hormonas de la pituitaria anterior, su regulación es la única en que la influencia predominante de su secreción hipotalámica es inhibitoria, en lugar de estimulante. Esta función inhibitoria se hace a través de la dopamina, neurotransmisor producido por el hipotálamo. El estímulo del pezón produce, por vía de un reflejo neuro-hormonal la liberación de prolactina. La prolactina se libera de la hipófisis anterior. El estímulo producido llega al hipotálamo, pasa a la hipófisis que libera prolactina la cual, vía sanguínea, alcanza las células del alvéolo mamario produciéndose la secreción de leche. Para que la leche pase de los alveolos a los conductos y se encuentre disponible para el lactante se requiere de la contracción de las células musculares que rodean al alvéolo, proceso que es producido por la oxitocina. El reflejo producido por la succión del pezón viaja por vía neurógena al hipotálamo estimulando la liberación de oxitocina desde la hipófisis posterior. La acción de la oxitocina se manifiesta por la sensación de plenitud mamaria y por un aumento del flujo sanguíneo; clínicamente puede observarse la secreción espontánea de leche del otro pecho mientras dura la succión. La eyección de la leche es fuertemente dependiente de la estimulación.
En las mujeres la principal acción de la prolactina es iniciar y mantener la lactancia.
La insuficiencia de prolactina no causa alteraciones en las mujeres que no están en período de lactancia y no causa anomalías en los hombres.Las altas concentraciones de prolactina resultan en una disminución de la secreción de gonadotropinas y por lo tanto, la disminución de la función ovárica. La frecuencia de los períodos menstruales disminuye o incluso pueden desaparecer por completo (amenorrea). En adición, se presentan síntomas de deficiencia de estrógenos, como la pérdida de deseo sexual, sequedad de la vagina, infertilidad, y con menor frecuencia, la lactancia anormal (galactorrea).En los hombres, las concentraciones elevadas de prolactina también disminuyen la secreción de gonadotropinas, y por lo tanto disminuyen la función testicular, lo que resulta en bajas concentraciones séricas de testosterona. En esos casos los principales síntomas son la pérdida del deseo sexual, disfunción eréctil, debilidad muscular e infertilidad.
Somatotrofina (GH).
A la síntesis y a la secreción de la GH hipofisaria las controla el hipotálamo mediante dos neurohormonas: una de carácter estimulador, la hormona liberadora de hormona de crecimiento (GHRH), y otra de carácter inhibidor, la somatostatina (SS o SRIF). La secreción es controlada casi por completo en respuesta estas hormonas secretadas en el hipotálamo y que son transportadas después a la para que actúen sobre las células somatotropas de la hipófisis anterior la GHRH y la hormona inhibidora de la secreción de la hormona del crecimiento o somatostatina o GHIH. La GH aumenta la entrada de aminoácidos en el interior de las células (en especial, las de músculo esquelético, hepatocitos y adipocitos) y, por tanto, aumenta la síntesis de proteínas (anabolismo proteico) en las células del organismo a la vez que reduce la desintegración o catabolismo de proteínas. Produce un aumento de la síntesis de DNA y de RNA y de la división celular. Debido a estos efectos, aumenta el crecimiento del esqueleto y de los músculos esqueléticos durante la niñez y la adolescencia. En adultos, ayuda a mantener el tamaño de huesos y músculos y promueve la reparación tisular. la GH estimula el catabolismo de la grasa almacenada en el tejido adiposo, con lo que aumenta la liberación de ácidos grasos libres al plasma que son aprovechados por las células del organismo para obtener energía al estimular su conversión a acetil-coenzima A.En general, su papel es promover el crecimiento de los órganos y tejidos y presenta un papel anabólico. Causa aumento de volumen de las células e incrementa las mitosis desarrollándose un número más elevado de células. Incrementa tanto la retención de calcio como la mineralización de los huesos. Induce la síntesis proteica. Disminuye la degradación proteica. Estimula el sistema inmunológico. Incrementa el transporte de la glucosa.
Hiposecreción: Si es congénita, o se produce durante la infancia, provoca enanismo. En general, las distintas partes del cuerpo conservan su proporción, pero el desarrollo total está muy disminuido. La pubertad sexual se presenta tan solo en un 30% de ellos. Si se presenta en el estado adulto (a causa de un tumor, o de una trombosis de los vasos hipofisiarios), se producen los siguientes efectos: hipotiroidismo, disminución de la producción de glucocorticoides, supresión de las gonadotropinas, hasta desaparecer la función sexual. Hipersecreción: En los casos en los que la adenohipófisis secreta exceso de GH durante los años de crecimiento del niño, los huesos crecen más rápido de lo normal produciéndose gigantismo. El gigante suele presentar hiperglucemia. Si el aumento de la secreción se produce después de la consolidación de los cartílagos de los huesos largos, la persona ya no puede crecer más, pero los tejidos blandos pueden ir aumentando de tamaño y los huesos aumentan de espesor. El trastorno recibe el nombre de acromegalia: el crecimiento es más notable en los huesos cortos de manos y pies, y en los huesos del cráneo, nariz, mandíbula y otros. La mandíbula se vuelve prominente, la nariz puede alcanzar el doble del tamaño normal, los huesos engruesan considerablemente y la mano se vuelve casi el doble más grande que antes. La frente se inclina hacia atrás, debido a que las órbitas de los ojos se desarrollan considerablemente. Crecen también tejidos blandos tales como lengua, hígado y sobre todo riñones.
Hormona melanocitoestimulante (MSH).
La hormona estimulante de los melanocitos es una hormona producida por la adenohipófisis o hipófisis anterior. Se ha demostrado que la α-MSH se une a un receptor (MC-1) que se encuentra en la membrana de los melanocitos humanos y que al ser estimulado da lugar a la activación de la tirosinasa, un enzima necesario para producir melanina. Los melanocitos tienen unas prolongaciones largas y delgadas que transfieren gránulos de melanina a los queratinocitos, en cuyo citoplasma son agrupados para formar un velo protector sobre su núcleo, evitando de este modo que el material genético sea dañado por la luz ultravioleta.
Hiposecreción: -Vitíligo (pérdida de pigmentación de la piel en ciertas áreas.) Albinismo (trastornos heredados que se caracterizan por producir poca o ninguna cantidad de melanina). -Piebaldismo
(enfermedad autosómico dominante que produce áreas de despigmentación debida a la ausencia de melanocitos en las zonas afectadas) -S. Waardenburg (se caracteriza por sordera congénita y defectos menores en estructuras derivadas de la cresta neural, que resultan en anomalías en la pigmentación de ojos, cabello y piel). Hipersecreción: -Lentigos (Manchas cutáneas planas de color tostado, marrón o negro)
-S. Peutz-Jegher (es una entidad rara autosómica dominante, caracterizada por lesiones hiperpigmentadas en boca, manos y pies; con presencia de pólipos gastrointestinales que ocasionan cuadros de anemia aguda o crónica, obstrucción intestinal y dolor abdominal)
Núcleos hipotalámicos
1 Anterior Procesamiento de emociones, mecanismos de la memoria reciente
2 Dorsomedial Integración de la información somática, visceral y olfativa y su relación con los sentimientos emocionales y los estados subjetivos
3 Lateral dorsal, Posterior, Pulvinar Desconocido
4 Ventral anterior Influye en la actividad de la corteza motora
5 Ventral lateral Influye en la actividad de la corteza motora
6 Posteromedial Relaciona con las sensaciones comunes con la consciencia
7 Posterolateral Relaciona con las sensaciones comunes con la consciencia
8 Intralaminar Influye en los niveles de consciencia y alerta
9 Línea media Desconocida
10 Reticular ¿La corteza cerebral regula al tálamo?
11 Cuerpo geniculado medial Procesos auditivos
12 Cuerpo geniculado lateral Información visual procedente del campo visual opuesto


{kind=link}
Regulación de la secreción hormonal
La mayoría de hormonas son liberadas en descargas cortas con poca o ninguna
secreción entre las descargas. Cuando es estimulada, una glándula endocrina
liberará su hormona en descargas más frecuentes y así el nivel en sangre de esta
hormona aumentará. En ausencia de estimulación, las descargas son mínimas o
están inhibidas y el nivel de hormona en sangre disminuye. La magnitud de la
secreción de cada hormona está regulada con mucha precisión de modo que se
evita el exceso o el defecto de su producción. La secreción hormonal por las
glándulas endocrinas es estimulada o inhibida por:
• Señales del sistema nervioso
• Cambios químicos en la sangre
• Otras hormonas.
Por ejemplo, los impulsos nerviosos a la médula adrenal regulan la liberación de
adrenalina, los niveles de calcio en sangre regulan la secreción de hormona
paratiroidea y la corticotropina (una hormona de la hipófisis anterior) estimula la
liberación de cortisol por la corteza suprarrenal.
En la mayor parte de los casos, la regulación de la secreción hormonal se ejerce
por un mecanismo de retroalimentación negativa (feedback negativo). Si la
hormona A aumenta la concentración en plasma del sustrato B, el aumento del
sustrato B inhibirá la secreción de la hormona A y la disminución del sustrato B
estimulará la secreción de la hormona A. Es decir que en el mecanismo de
retroalimentación negativa, la respuesta producida por la hormona en el órgano
diana, tiene un efecto inhibidor sobre el estímulo inicial. Esta relación puede
existir entre una hormona y uno o más sustratos, niveles de minerales en plasma,
otras hormonas u otros factores como el volumen extracelular.
Ocasionalmente un mecanismo de retroalimentación positiva (feedback
positivo) contribuye a la regulación de la secreción hormonal. Un ejemplo ocurre
durante el parto. La oxitocina estimula las contracciones del útero. A su vez las
contracciones del útero estimulan más liberación de oxitocina. Es decir que en el
mecanismo de retroalimentación positiva, la respuesta producida por la hormona
intensifica el estímulo inicial.
Algunos patrones de regulación de secreción hormonal siguen los ciclos
ambientales como la luz/oscuridad o el sueño/vigilia. Los ritmos de 24 horas se
conocen como ritmos circadianos. La secreción de diversas hormonas como la
corticotropina (ACTH), el cortisol, la hormona del crecimiento y la prolactina
siguen ritmos circadianos. El conocer estos patrones de secreción es importante
para interpretar los resultados de los análisis realizados en muestras de sangre
obtenidas en diversos momentos del día.
secreción entre las descargas. Cuando es estimulada, una glándula endocrina
liberará su hormona en descargas más frecuentes y así el nivel en sangre de esta
hormona aumentará. En ausencia de estimulación, las descargas son mínimas o
están inhibidas y el nivel de hormona en sangre disminuye. La magnitud de la
secreción de cada hormona está regulada con mucha precisión de modo que se
evita el exceso o el defecto de su producción. La secreción hormonal por las
glándulas endocrinas es estimulada o inhibida por:
• Señales del sistema nervioso
• Cambios químicos en la sangre
• Otras hormonas.
Por ejemplo, los impulsos nerviosos a la médula adrenal regulan la liberación de
adrenalina, los niveles de calcio en sangre regulan la secreción de hormona
paratiroidea y la corticotropina (una hormona de la hipófisis anterior) estimula la
liberación de cortisol por la corteza suprarrenal.
En la mayor parte de los casos, la regulación de la secreción hormonal se ejerce
por un mecanismo de retroalimentación negativa (feedback negativo). Si la
hormona A aumenta la concentración en plasma del sustrato B, el aumento del
sustrato B inhibirá la secreción de la hormona A y la disminución del sustrato B
estimulará la secreción de la hormona A. Es decir que en el mecanismo de
retroalimentación negativa, la respuesta producida por la hormona en el órgano
diana, tiene un efecto inhibidor sobre el estímulo inicial. Esta relación puede
existir entre una hormona y uno o más sustratos, niveles de minerales en plasma,
otras hormonas u otros factores como el volumen extracelular.
Ocasionalmente un mecanismo de retroalimentación positiva (feedback
positivo) contribuye a la regulación de la secreción hormonal. Un ejemplo ocurre
durante el parto. La oxitocina estimula las contracciones del útero. A su vez las
contracciones del útero estimulan más liberación de oxitocina. Es decir que en el
mecanismo de retroalimentación positiva, la respuesta producida por la hormona
intensifica el estímulo inicial.
Algunos patrones de regulación de secreción hormonal siguen los ciclos
ambientales como la luz/oscuridad o el sueño/vigilia. Los ritmos de 24 horas se
conocen como ritmos circadianos. La secreción de diversas hormonas como la
corticotropina (ACTH), el cortisol, la hormona del crecimiento y la prolactina
siguen ritmos circadianos. El conocer estos patrones de secreción es importante
para interpretar los resultados de los análisis realizados en muestras de sangre
obtenidas en diversos momentos del día.
Acción de las hormonas
La respuesta celular a una hormona depende tanto de la hormona como de la célula diana. Varias células diana responden de un modo diferente a la misma hormona. La insulina, por ejemplo, estimula la síntesis de glucógeno en las células hepáticas y la síntesis de triglicéridos en los adipocitos. Con frecuencia, la respuesta a una hormona es la síntesis de nuevas moléculas. Otros efectos hormonales son: producir cambios en la permeabilidad de la membrana de la célula diana, estimular el transporte de una sustancia dentro o fuera de la célula diana, alterar la velocidad de reacciones metabólicas específicas o causar la contracción del músculo liso o cardíaco. En parte, estos efectos variados de las hormonas son posibles debido a que hay varios mecanismos diferentes de acción hormonal.
Las hormonas, casi de modo invariable, se combinan primero con receptores
hormonales situados en la superficie o en el interior de las células diana. Una célula puede tener simultáneamente receptores en la membrana celular y en el citoplasma. Asimismo una célula puede disponer de diversos receptores para un tipo de hormona, por ejemplo, varios receptores de membrana para diversas hormonas peptídeas. La combinación de hormona y receptor suele iniciar una cascada de reacciones en la célula. Cada receptor suele ser muy específico para una hormona determinada. Los tejidos diana que se ven afectados por una hormona son los que contienen los receptores específicos para esta hormona.
ACTIVACIÓN DE RECEPTORES INTRACELULARES
Las hormonas esteroides y las tiroideas (tiroxina y triyodotironina) pasan fácilmente a través de las membranas plasmáticas porque son liposolubles. Una vez que ha entrado en la célula, la hormona se une a/ y activa un receptor intracelular. En el caso de las hormonas esteroides, sus receptores están en el citoplasma, son receptores citoplasmáticos, y una vez la hormona se une a su receptor, el complejo hormona-receptor penetra en el núcleo y actúa sobre la expresión genética, es decir, se ponen en marcha o se detienen genes específicos del ADN nuclear. Cuando el ADN es trascrito, nuevas formas de ARN mensajero dejan el núcleo y entran en el citoplasma. Allí dirigen la síntesis de nuevas proteínas, usualmente enzimas, en los ribosomas, que causan las respuestas fisiológicas que son características de esa hormona. Hay que señalar que el complejo hormona-receptor puede tener efectos ya en el citoplasma, independientes de los efectos producidos en el núcleo celular. En el caso de las hormonas tiroideas, sus receptores están en el núcleo, son
receptores nucleares que se unen al ADN en la región promotora de genes regulados por dichas hormonas. De modo que cuando las hormonas tiroideas entran en el núcleo, se unen a sus receptores y promueven la trascripción de un gran número de genes codificadores de un amplio rango de proteínas.
ACTIVACIÓN DE RECEPTORES DE MEMBRANA PLASMÁTICA
La adrenalina, noradrenalina, péptidos y proteínas no son liposolubles y, por tanto, no pueden pasar a través de la membrana celular. Los receptores de estas hormonas hidrosolubles se encuentran en la superficie externa de la membrana plasmática. Ya que cada una de estas hormonas solo puede dar su mensaje a la
membrana plasmática, se la llama primer mensajero. Pero se necesita un segundo mensajero para trasladar el mensaje dentro de la célula donde tienen lugar las respuestas hormonales. Hay diversos segundos mensajeros como el AMP cíclico, el calcio o el inositol trifosfato. Una hormona puede usar más de un segundo mensajero. El segundo mensajero mejor conocido es el AMP cíclico (AMPc). Al unirse una
hormona (primer mensajero) a su receptor de membrana, se activan proteínas reguladoras unidas a la membrana, las proteínas G, que, a su vez, activan moléculas de adenil ciclasa, enzima situado en la superficie interna de la membrana que entonces sintetiza AMP cíclico a partir del ATP en el citoplasma celular. El AMP cíclico actúa como segundo mensajero pero no produce directamente una respuesta fisiológica. Lo que hace es activar uno o más enzimas llamados colectivamente proteínas quinasas que pueden estar libres en el citoplasma o unidos a la membrana plasmática. Las proteínas quinasas son enzimas fosforiladores, lo que significa que extraen un grupo fosfato del ATP y lo añaden a una proteína, que suele ser un enzima. La fosforilación activa unos enzimas e inactiva otros. El resultado de fosforilar un enzima particular puede ser la
regulación de otros enzimas, la síntesis de proteínas o el cambio en la permeabilidad de la membrana plasmática, entre otros efectos. Existen diferentes proteína qinasas dentro de diferentes células diana y dentro de diferentes organelas de la misma célula. Así, una proteína quinasa podría estar involucrada en la síntesis de glucógeno, otra en el catabolismo de lípidos, otra en la síntesis proteica etc. Por ejemplo, la elevación de AMP cíclico provoca que en los adipocitos se rompan los triglicéridos y se liberen ácidos grasos más rápidamente. Tras un breve período de tiempo, un enzima llamado fosfodiestarasa inactiva el AMP cíclico. De este modo la respuesta celular termina hasta que nueva hormona se una a los
receptores de la membrana. Las hormonas que se unen a receptores de la membrana pueden inducir sus efectos a muy bajas concentraciones porque inician una cascada, o reacción en cadena, de efectos. Cada paso en la cadena multiplica o amplifica el efecto inicial. Es lo que se llama amplificación de los efectos hormonales. Por ejemplo, cuando una sola molécula de adrenalina se une a su receptor en un hepatocito, puede activar unas cien moléculas de proteína G. A su vez, cada proteína G activa una molécula de adenil ciclasa. Si cada adenil ciclasa produce unos mil AMP cíclicos, entonces 100000 de estos segundos mensajeros serán liberados dentro de la célula. Cada AMP cíclico puede activar una proteína quinasa que, a su vez, puede actuar sobre cientos o miles de moléculas de sustrato. Algunas de las quinasas fosforilan y activan un enzima clave para el catabolismo del glucógeno. El resultado final de la unión de la adrenalina a su receptor en un hepatocito, es la ruptura de millones de moléculas de glucógeno a glucosa.
INTERACCIONES HORMONALES
La respuesta de una célula diana a una hormona depende de la concentración de la hormona y del número de receptores. Pero también es importante el modo en que las hormonas interaccionan con otras hormonas. Hay varios tipos de interacción:
• Efecto permisivo, el efecto de una hormona sobre una célula diana requiere una exposición previa o simultánea a otra u otras hormonas. Por ejemplo, un aumento de estrógenos puede dar lugar a un aumento en el número de receptores de progesterona. Ambas hormonas preparan el útero para la posible implantación de un zigoto o huevo fertilizado• Efecto sinérgico, dos o más hormonas complementan sus respectivas acciones y ambas son necesarias para conseguir la respuesta hormonal total. Por ejemplo, la producción, secreción y salida de leche por las glándulas mamarias requieren el efecto sinérgico de estrógenos, progesterona, prolactina y oxitocina
• Efecto antagonista, el efecto de una hormona sobre una célula diana es contrarrestado por otra hormona. Un ejemplo es la insulina que desciende los niveles de glucosa en sangre y el glucagón, que hace lo contrario.
jueves, 15 de marzo de 2018
Hormonas adenohipófisis
La hipófisis consta de tres partes: adenohipófisis,lóbulo intermedio (pars intermedia) y neurohipófisis.
Mediante distintas técnicas se lograron aislar diferente grupos celulares:
1. Células corticotrofas. Producen hormonas corticotrofas (ACTH).
2. Células gonadotrofas. Sintetizan y segregan gonadotropinas. (FSH, LH), activinas e inhibinas.
3. Células lactotrofas: sintetizan y segregan prolactina (PRL). Su proximidad a las células gonadotrofas sugiere actividad parácrina relacionadas.
4. Células melanotróficas: sintetizan y segregan hormonas melanotropas (MSH).
5. Células somatotrofas: sintetiza y segrega hormona de crecimiento (GH).
6. Células tirotrofas: sintetizan y segregan tirotrofina (TSH).
7. Células cromófobas: no secretoras, agranulares o nulas (Null cells). No sintetizan hormonas.
Hormona del crecimiento humano (hGH) o somatotropina. Cuyo órgano diana principal es el hígado. Estimula la síntesis y secreción de factores de crecimiento insulinoides (FCI) en el hígado, músculos, cartílago, huesos y otros tejidos; los FCI fomentan el crecimiento de las células corporales, síntesis de proteínas, reparación de tejidos, lipólisis y aumento de glucemia.En general, su papel es promover el crecimiento de los órganos y tejidos y presenta un papel anabólico. Causa aumento de volumen de las células e incrementa las mitosis desarrollándose un número más elevado de células. Incrementa tanto la retención de calcio como la mineralización de los huesos. Induce la síntesis proteica.Disminuye la degradación proteica.Estimula el sistema inmunológico.Incrementa el transporte de la glucosa.
Hormona estimulante de la tiroides (TSH) o tirotropina. Cuyo órgano diana principal es la tiroides. Estimula la síntesis y secreción de hormonas tiroideas. Las hormonas T4 y T3 que fabrica la glándula tiroides participan en el desarrollo del ser humano e intervienen en todas las etapas de la vida. Durante la infancia estimulan el desarrollo, el crecimiento y la maduración del sistema nervioso central, mientras que en la vida adulta se encargan de regular el metabolismo de todos los órganos y sistemas. Estas hormonas controlan el nivel del calcio sanguíneo y el del colesterol, el ritmo cardíaco, la frecuencia cardíaca, la presión arterial, la temperatura de nuestro cuerpo y la sudoración. Asimismo, juegan un papel importante en la producción de glucógeno y en la utilización de la glucosa, en la contracción de los músculos, en el movimiento de los intestinos y en la fertilidad de la mujer. Otras funciones en las que tienen un papel determinante son el control de la temperatura del cuerpo, la memoria y el metabolismo, la digestión y la formación de vitamina A a partir de los carotenos que aportan los alimentos.
Hipófisis anterior (adenohipófisis).
Mediante distintas técnicas se lograron aislar diferente grupos celulares:1. Células corticotrofas. Producen hormonas corticotrofas (ACTH).
2. Células gonadotrofas. Sintetizan y segregan gonadotropinas. (FSH, LH), activinas e inhibinas.
3. Células lactotrofas: sintetizan y segregan prolactina (PRL). Su proximidad a las células gonadotrofas sugiere actividad parácrina relacionadas.
4. Células melanotróficas: sintetizan y segregan hormonas melanotropas (MSH).
5. Células somatotrofas: sintetiza y segrega hormona de crecimiento (GH).
6. Células tirotrofas: sintetizan y segregan tirotrofina (TSH).
7. Células cromófobas: no secretoras, agranulares o nulas (Null cells). No sintetizan hormonas.
Hormona del crecimiento humano (hGH) o somatotropina. Cuyo órgano diana principal es el hígado. Estimula la síntesis y secreción de factores de crecimiento insulinoides (FCI) en el hígado, músculos, cartílago, huesos y otros tejidos; los FCI fomentan el crecimiento de las células corporales, síntesis de proteínas, reparación de tejidos, lipólisis y aumento de glucemia.En general, su papel es promover el crecimiento de los órganos y tejidos y presenta un papel anabólico. Causa aumento de volumen de las células e incrementa las mitosis desarrollándose un número más elevado de células. Incrementa tanto la retención de calcio como la mineralización de los huesos. Induce la síntesis proteica.Disminuye la degradación proteica.Estimula el sistema inmunológico.Incrementa el transporte de la glucosa.
Hormona estimulante de la tiroides (TSH) o tirotropina. Cuyo órgano diana principal es la tiroides. Estimula la síntesis y secreción de hormonas tiroideas. Las hormonas T4 y T3 que fabrica la glándula tiroides participan en el desarrollo del ser humano e intervienen en todas las etapas de la vida. Durante la infancia estimulan el desarrollo, el crecimiento y la maduración del sistema nervioso central, mientras que en la vida adulta se encargan de regular el metabolismo de todos los órganos y sistemas. Estas hormonas controlan el nivel del calcio sanguíneo y el del colesterol, el ritmo cardíaco, la frecuencia cardíaca, la presión arterial, la temperatura de nuestro cuerpo y la sudoración. Asimismo, juegan un papel importante en la producción de glucógeno y en la utilización de la glucosa, en la contracción de los músculos, en el movimiento de los intestinos y en la fertilidad de la mujer. Otras funciones en las que tienen un papel determinante son el control de la temperatura del cuerpo, la memoria y el metabolismo, la digestión y la formación de vitamina A a partir de los carotenos que aportan los alimentos.
Hormona foliculoestimulante (FSH). Cuyos órganos diana son los ovarios y testículos. En las mujeres: inicia el desarrollo de los ovocitos e induce la secreción ovárica de estrógenos. En los hombres: estimula la producción de espermatozoides en testículos. La FSH, en la hembra, actúa sobre los folículos
en los que se encuentran los óvulos en desarrollo, produciendo
su crecimiento además de iniciar la secreción de la hormona
sexual femenina, el estrógeno, que al alcanzar determinados niveles, inhibe la secreción hipofisiaria de la FSH. Durante la fase folicular, la FSH estimula la producción de estrógenos en células de la granulosa, al estimular la conversión de andrógenos en estrógenos. La FSH estimula las células de la granulosa y de Sertoli para que estas excreten estrógenos. Uniéndose a sus receptores en la membrana de estas células y actuando el AMP-c como segundo mensajero, la FSH aumenta la transcripción del gen de la aromatasa, la enzima específica de la síntesis del estradiol. La FSH también incrementa el número de receptores a la LH en las células diana, aumentando por tanto su sensibilidad a esta hormona. Finalmente, estimula la producción de inhibina y otros péptidos en las células de la granulosa y de Sertoli. En el macho, esta hormona promueve la espermatogénesis.
en los que se encuentran los óvulos en desarrollo, produciendo
su crecimiento además de iniciar la secreción de la hormona
sexual femenina, el estrógeno, que al alcanzar determinados niveles, inhibe la secreción hipofisiaria de la FSH. Durante la fase folicular, la FSH estimula la producción de estrógenos en células de la granulosa, al estimular la conversión de andrógenos en estrógenos. La FSH estimula las células de la granulosa y de Sertoli para que estas excreten estrógenos. Uniéndose a sus receptores en la membrana de estas células y actuando el AMP-c como segundo mensajero, la FSH aumenta la transcripción del gen de la aromatasa, la enzima específica de la síntesis del estradiol. La FSH también incrementa el número de receptores a la LH en las células diana, aumentando por tanto su sensibilidad a esta hormona. Finalmente, estimula la producción de inhibina y otros péptidos en las células de la granulosa y de Sertoli. En el macho, esta hormona promueve la espermatogénesis.
Hormona luteinizante (LH). Cuyos órganos diana principales son ovarios y testículos. En las mujeres: estimula la secreción de estrógenos y progesterona, ovulación y formación del cuerpo lúteo. En los hombres: estimula el desarrollo de las células intersticiales de Leydig en los testículos y su producción de testosterona.En las mujeres, un aumento de la LH desencadena la ovulación. Influye en el incremento de la actividad aromatasa en las células de la granulosa y en la producción de andrógenos en las células de la teca durante la maduración folicular preovulatoria. En el momento de la maduración del folículo, el aumento de estrógenos conduce a una liberación de LH que desencadena la ovulación, no sólo liberando el óvulo sino también iniciando la conversión de los folículos residuales en un cuerpo lúteo que, a su vez, produce progesterona con el fin de preparar al endometrio para una posible implantación. En el hombre, la LH actúa sobre las células de Leydig del testículo y se encarga de la producción de testosterona, un andrógeno que ejerce actividad endocrina y actividad intratesticular (como la
espermatogénesis).
espermatogénesis).
Prolactina (PRL). Cuyo órgano diana principal son las glándulas
mamarias. Aunada a otras, estimula la secreción de leche en las glándulas mamarias.En las mujeres la principal acción de la prolactina es iniciar y mantener la lactancia.
Adrenocorticotropina (ACTH) o corticotropina. En la corteza suprarrenal estimula la secreción de glucocorticoides (principalmente cortisol). Glucocorticoides sobre el metabolismo de los hidratos de carbono es el aumento de la producción de glucosa mediante el aumento de la gluconeogénesis hepática.También aumentan la resistencia celular a la insulina, por lo que disminuyen la entrada de glucosa a la célula.El MC más importante es la aldosterona y en menor grado la 11-desoxicorticosterona. Los MC tienen acciones más limitadas que los GCC. Su función fundamental es mantener el volumen intravascular conservando sodio y eliminando potasio e hidrogeniones. Ejercen estas acciones en el riñón, el intestino y las glándulas salivales y sudoríparas. Muchas acciones de los andrógenos suprarrenales se ejercen mediante su conversión a andrógenos o estrógenos activos, como la testosterona, la dihidrotestosterona, la estrona y el estradiol.
Hormona estimulante de los melanocitos (MSH). Puede estar relacionada con el oscurecimiento de la piel.
Hormonas hipotalámicas
Las hormonas hipotalámicas son péptidos de pequeño tamaño con actividad fisiológica exclusiva en concentraciones elevadas observables en el sistema porta hipofisiario, con la excepción de los péptidos conocidos como hormonas de la neurohipófisis: la hormona antidiurética (ADH, arginina-vasopresina) y la oxitocina.
Las células en el hipotálamo sintetizan al menos 9 hormonas distintas, y la glándula hipófisis 7. Juntas estas 16 hormonas juegan papeles importantes en la regulación de aspectos del crecimiento, el desarrollo, el metabolismoy la homeostasis. La relación está dada en que el lóbulo anterior de la hipófisis (adenohipófisis) es controlado por el hipotálamo mediante la segregación de sustancias parecidas a las hormonas, que llegan hasta los vasos sanguíneos que conectan a las dos zonas; y el lóbulo posterior (neurohipófisis) igualmente es controlado por el hipotálamo mediante impulsos nerviosos, es decir que, por un lado, el hipotálamo conecta con la hipófisis anterior (adenohipófisis) y por otro lado, el hipotálamo conecta con la hipófisis posterior (neurohipófisis).
Entre estas hormonas secretadas a nivel hipotalámico hacia adenohipófisis comprenden:
1. Hormonas estimulantes:
Las células en el hipotálamo sintetizan al menos 9 hormonas distintas, y la glándula hipófisis 7. Juntas estas 16 hormonas juegan papeles importantes en la regulación de aspectos del crecimiento, el desarrollo, el metabolismoy la homeostasis. La relación está dada en que el lóbulo anterior de la hipófisis (adenohipófisis) es controlado por el hipotálamo mediante la segregación de sustancias parecidas a las hormonas, que llegan hasta los vasos sanguíneos que conectan a las dos zonas; y el lóbulo posterior (neurohipófisis) igualmente es controlado por el hipotálamo mediante impulsos nerviosos, es decir que, por un lado, el hipotálamo conecta con la hipófisis anterior (adenohipófisis) y por otro lado, el hipotálamo conecta con la hipófisis posterior (neurohipófisis).
Entre estas hormonas secretadas a nivel hipotalámico hacia adenohipófisis comprenden:
1. Hormonas estimulantes:
- Hormona liberadora de corticotropina (CRH) .La CRH es activadora de la secreción hipofisiaria de ACTH (producto derivado del gen determinante de la propiomelanocortina, POMPC). Su secreción procede de neuronas hipotalámicas del núcleo paraventricular, núcleo supraóptico, núcleo arcuato y sistema límbico. Actúa fijándose a receptores específicos de las células corticotropicas y solo estimula la liberación hormonal en presencia de calcio. La concentración de AMPc aumenta paralelamente al efecto biológico, aumentando la concentración de RNA m de la POMC. El efecto estimulante de la CRH sobre la producción del AMPc es disminuido por los glucocorticoides.
- Hormona liberadora de la hormona de crecimiento (GHRH). El núcleo arqueado del hipotálamo es el lugar principal de producción de GHRH, aunque también existen algunas neuronas de GHRH en el núcleo ventromedial. Después de fijarse en la membrana de las células hipofisarias estimula la secreción de GH por un mecanismo dependiente de calcio y activa a la adenilciclasa, con la acumulación delAMPc. También activa el ciclo del fosfatidilinositol y ejercería una acción directa dentro de la célulamediante fosforilación de una enzima ligada al gránulo secretorio. La GHRH también aumenta la formaciónde nueva GH estimulando la trascripción del RNAm específico para GH, los efectos de la GHRH son bloqueados por la somatostatina y potenciados por los glucocorticoides.
- Hormona liberadora de gonadotropina (GnRH ó LHRH)
La GnRH actúa sobre los receptores hipofisarios de alta afinidad para estimular la producción y liberación de FSH y LH. Al igual que otros neuropéptidos se sintetiza como parte de una gran prohormona que se cliva enzimáticamente y es modificada todavía más dentro de los gránulos secretorios; Se secreta principalmente por neuronas del área preóptica y está constituida por tan solo 10 aminoácidos. La acción de la LHRH sobre la hipófisis se inicia con la fijación a receptores específicos de la superficie celular, el proceso de liberación se activa mediante la movilización del calcio intracelular. Los agonistas adrenérgicos facilitan aparentemente la liberación de LHRH, mientras que los opiáceos endógenos la inhiben, los estrógenos aumentan la cantidad de receptores de LHRH y los andrógenos la reducen. La respuesta hipofisaria a la LHRH varía considerablemente a lo largo de la vida. La LHRH y lasgonadotrofinas aparecen inicialmente en el feto, aproximadamente a las 10 semanas de gestación. La LHRH desencadena un aumento brusco de gonadotrofinas en los 3 primeros meses después del parto. Posteriormente, la sensibilidad de la LHRH disminuye hasta el inicio de la pubertad. La respuesta de la FSH a la LHRH es mayor que la de la LH antes de la pubertad. Sin embargo, con la pubertad aumenta la sensibilidad para la LHRH y comienza la secreción pulsátil de LH, inicialmente durante el sueño. Más adelante y durante los años de vida fértil, estas pulsaciones aparecen a lo largo del día y la respuesta de LH es mayor que la de FSH. Tras la menopausia, se elevan los valores de las gonadotrofinas, aumentando más los de FSH que los de LH.
- Hormona liberadora de tirotropina (TSHRH)
Es un tripéptido, se produce en el área hipotalámica anterior, también se ha encontrado TSHRH
extrahipotalámica en la hipófisis posterior, en otras zonas del cerebro y de la médula espinal y en el aparato
gastrointestinal. La TSHRH estimula la secreción de TSH mediante el incremento del calcio citoplasmático
libre, probablemente el fosfatidilinositol y los fosfolípidos de membrana participan en la secreción de TSH
mediada por la TSHRH y también estimula la liberación de prolactina. Los efectos estimulantes de la TSHRH se inician con la fijación del péptido a los receptores específicos en la membrana plasmática de la célula hipofisaria. La acción de la TSHRH se ejerce sobre la membrana y no depende de la internalización, aunque esta última tiene lugar. Al principio se pensó que la TSHRH actuaba activando a la adenilciclasa de membrana con la formación de AMPc. Un mecanismo alternativo (o complementario) sería una hidrólisis calcio dependiente del fosfatidilinositol, con fosforilación de proteínquinasas claves como paso crucial en la activación posreceptor. La TSHRH, estimula la formación de RNAm que codifica a la prolactina.
- Factores liberadores de prolactina (PRL)
Los factores liberadores de PRL son neurotransmisores (serotonina, acetilcolina), sustancias opiáceas y
estrógenos.
Otros factores estimulantes de la liberación de PRL en la especie humana son la TSHRH, péptido intestinal
vasoactivo(VIP), la sustancia P, colecistoquinina, neurotensina, GHRH, GnRH, oxitocina, vasopresina y
galanina. Estos factores se han observado experimentalmente, si bien su significado fisiológico en la especie
humana es desconocido.
2.Hormonas inhibidoras
Factores inhibidores de PRL
Desde el punto de vista fisiológico se ha comprobado como la dopamina (DA) constituye el principal factor
hipotalámico con actividad inhibidora de la liberación de PRL. Los núcleos arqueados y paraventricular del
hipotálamo producen dopamina: la DA viaja a través de los axones hasta las terminaciones nerviosas de la
eminencia media, donde se libera (sistema de la DA tuberoinfundibular) a la circulación portal y llega a la
hipófisis anterior para inhibir la liberación de PRL a través de las interacciones con los receptores D2
(receptores de la DA ligados a la adenilatociclasa). La DA inhibe la formación de AMPc e inhibe la síntesis de fosfoinisitol, paso importante en la regulación posreceptor de la secreción de PRL.
Hormona inhibidora de GH (GHRIH o somatostatina)
Es una hormona de 14 aminoácidos que se sintetiza como una pre-prohormona de 116 aa. Y una pre-hormona de 28 aa., y se halla ampliamente distribuida en todas las células del sistema nervioso, actuando comoneurotransmisor en muchas regiones como la médula espinal, el tronco encefálico y la corteza cerebral. Lasomatostatina también existe en el aparato gastrointestinal y en otros órganos. Las células específicas
secretoras de somatostatina (células D) de los islotes pancreáticos intervienen en la regulación de la secreciónde insulina y glucagón y constituyen un ejemplo de control parácrino hormonal. La somatostatina actúa a través de una familia de al menos cinco receptores de membranas acoplados a proteína G que utilizan diversas vías de segundos mensajeros. La somatostatina inhibe la secreción de GH y reduce la respuesta de GH a los estímulos secretagogos sin alterar los niveles de RNMm de la GH.
La pre-hormona de 28 aa. Tiene una vida media más larga y es un inhibidor más potente de GH y de insulina.La somatostatina posee mayor afinidad por los receptores hipotalámicos y corticales e inhibe de manera más potente la liberación de glucagón, el flujo sanguíneo esplácnico, la motilidad intestinal y la secreción exócrina gástrica
Eje hipotalámico-hipofisiario
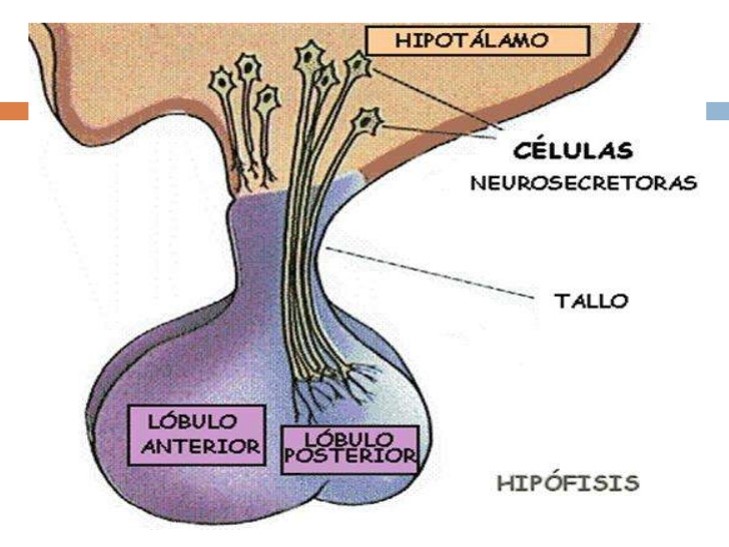
El eje hipotalámico-hipofisiario se refiere a la relación anatómica y funcional que existe entre las neuronas hipotalámicas, por una parte, y la adenohipófisis y la neurohipófisis, por otra.
El hipotálamo es un centro regulador importante del sistema nervioso así como una glándula endócrina crucial.Es una estructura del sistema nervioso central, ubicada a nivel del diencéfalo por debajo del tálamo. Está ampliamente inervado e irrigado. Consta de 10 núcleos por medio de los cuales ejerce sus funciones neuroendocrinas.


La comunicación entre la hipófisis anterior y el hipotálamo se efectúa a través de pequeños vasos
sanguíneos que proceden del hipotálamo y van a desembocar en los sinusoides (tipo especial de capilares) hipofisarios, proporcionando una conexión vascular directa entre el hipotálamo y las células endocrinas de la hipófisis anterior. Estos vasos de comunicación entre hipotálamo y adenohipófisis constituyen el sistema portal hipotálamo-hipofisario. De este modo, las hormonas liberadoras e inhibidoras del hipotálamo pasan a los capilares hipotalámicos y son transportadas por la sangre directamente a los sinusoides de la hipófisis anterior desde donde se ponen en contacto con los distintos tipos de células de la adenohipófisis para facilitar o inhibir su función secretora.
Cada tipo de hormona adenohipofisaria tiene su correspondiente hormona hipotalámica de liberación y algunas tienen también la correspondiente hormona hipotalámica de inhibición.
Las hormonas hipotalámicas se sintetizan en los cuerpos neuronales y son transportadas por los axones hasta la porción basal del hipotálamo, conocida como eminencia media. Desde allí, son liberadas a una red de capilares que confluyen en vénulas que se dirigen hacia la adenohipófisis y dan origen a una segunda red de capilares en la propia adenohipófisis. Este sistema vascular, conocido como sistema porta hipotálamo-hipofisario explica por qué las hormonas hipotalámicas, aún siendo secretadas en cantidades muy bajas, llevan a cabo su función sobre la adenohipófisis de una forma eficiente, ya que no llegan a alcanzar a la circulación sistémica y por tanto no se diluyen en el sistema vascular.
Mientras la conexión funcional con el lóbulo anterior de la hipófisis tiene lugar a través de un nexo vascular representado por el sistema porta-hipofisario que conduce las hormonas hipotalámicas liberadoras e inhibidoras, la neurohipófisis recibe directamente las prolongaciones axónicas de los núcleos supraóptico y paraventricular que sintetizan la oxitocina y la vasopresina. Existen numerosos mecanismos de retroalimentación específicos entre las hormonas hipofisarias, periféricas e hipotalámicas.El conjunto hipotálamo-hipófisis forman una unidad funcional capaz de modificar su producción hormonal para adaptarse a las necesidades del resto del organismo.
El conjunto hipotálamo-hipófisis forman una unidad funcional capaz de modificar su producción hormonal para adaptarse a las necesidades del resto del organismo. En general, el conjunto hipotálamo-hipófisis recibe
información del exterior a través del SN y del interior a través de la circulación sanguínea.Sangre arterial no llega directamente a la adenohipófisis. Ramas de la carótida interna llegan a la eminencia media, recoge hormonas producidas en el hipotálamo (sistema neuro-hipofisario) y las transporta junto con otras hormonas y sustancias periféricas hasta la hipófisis anterior. La hipófisis posterior, como el hipotálamo, también recibe sangre arterial directamente de ramas de la carótida interna. Por el contrario, la adenohipófisis sólo recibe sangre la arterial que previamente a pasado por la eminencia media o la neurohipófisis a través de los sistemas portahipofisarios largo y corto respectivamente. El retorno venoso de ambos lóbulos hipofisarios se hace a través de seno cavernoso.


El hipotálamo es un centro regulador importante del sistema nervioso así como una glándula endócrina crucial.Es una estructura del sistema nervioso central, ubicada a nivel del diencéfalo por debajo del tálamo. Está ampliamente inervado e irrigado. Consta de 10 núcleos por medio de los cuales ejerce sus funciones neuroendocrinas.
La glándula hipofisis tiene una forma de guisante que mide 1.5 cm aprox, y descansa en la silla turca del hueso esfenoides. Está unida al hipotálamo mediante un tallo y tiene dos lóbulos anatómica y funcionalemente separados:
- El lóbulo anterior (adenohipófisis), que secreta hormonas que regulan un amplio rango de actividades corporales. La liberación de las hormonas de esta región se estimula por hormonas liberadoras y se inhibe por hormonas inhibidoras desde el hipotálamo.

- El lóbulo posterior (neurohipófisis). La neurohipófisis contiene axones y terminales axónicos de más de 10000 neuronas cuyos cuerpos celulares se localizan en los núcleos supraóptico y paraventriculardel hipotálamo.

La comunicación entre la hipófisis anterior y el hipotálamo se efectúa a través de pequeños vasos
sanguíneos que proceden del hipotálamo y van a desembocar en los sinusoides (tipo especial de capilares) hipofisarios, proporcionando una conexión vascular directa entre el hipotálamo y las células endocrinas de la hipófisis anterior. Estos vasos de comunicación entre hipotálamo y adenohipófisis constituyen el sistema portal hipotálamo-hipofisario. De este modo, las hormonas liberadoras e inhibidoras del hipotálamo pasan a los capilares hipotalámicos y son transportadas por la sangre directamente a los sinusoides de la hipófisis anterior desde donde se ponen en contacto con los distintos tipos de células de la adenohipófisis para facilitar o inhibir su función secretora.
Cada tipo de hormona adenohipofisaria tiene su correspondiente hormona hipotalámica de liberación y algunas tienen también la correspondiente hormona hipotalámica de inhibición.
Las hormonas hipotalámicas se sintetizan en los cuerpos neuronales y son transportadas por los axones hasta la porción basal del hipotálamo, conocida como eminencia media. Desde allí, son liberadas a una red de capilares que confluyen en vénulas que se dirigen hacia la adenohipófisis y dan origen a una segunda red de capilares en la propia adenohipófisis. Este sistema vascular, conocido como sistema porta hipotálamo-hipofisario explica por qué las hormonas hipotalámicas, aún siendo secretadas en cantidades muy bajas, llevan a cabo su función sobre la adenohipófisis de una forma eficiente, ya que no llegan a alcanzar a la circulación sistémica y por tanto no se diluyen en el sistema vascular.
Mientras la conexión funcional con el lóbulo anterior de la hipófisis tiene lugar a través de un nexo vascular representado por el sistema porta-hipofisario que conduce las hormonas hipotalámicas liberadoras e inhibidoras, la neurohipófisis recibe directamente las prolongaciones axónicas de los núcleos supraóptico y paraventricular que sintetizan la oxitocina y la vasopresina. Existen numerosos mecanismos de retroalimentación específicos entre las hormonas hipofisarias, periféricas e hipotalámicas.El conjunto hipotálamo-hipófisis forman una unidad funcional capaz de modificar su producción hormonal para adaptarse a las necesidades del resto del organismo.
El conjunto hipotálamo-hipófisis forman una unidad funcional capaz de modificar su producción hormonal para adaptarse a las necesidades del resto del organismo. En general, el conjunto hipotálamo-hipófisis recibe
información del exterior a través del SN y del interior a través de la circulación sanguínea.Sangre arterial no llega directamente a la adenohipófisis. Ramas de la carótida interna llegan a la eminencia media, recoge hormonas producidas en el hipotálamo (sistema neuro-hipofisario) y las transporta junto con otras hormonas y sustancias periféricas hasta la hipófisis anterior. La hipófisis posterior, como el hipotálamo, también recibe sangre arterial directamente de ramas de la carótida interna. Por el contrario, la adenohipófisis sólo recibe sangre la arterial que previamente a pasado por la eminencia media o la neurohipófisis a través de los sistemas portahipofisarios largo y corto respectivamente. El retorno venoso de ambos lóbulos hipofisarios se hace a través de seno cavernoso.

Suscribirse a:
Comentarios (Atom)